
Functional Analysis At Structure Level
Prosite and Motif Scan are used for functional analysis at the sequence level for structure. For structure, it is seen that how the functional motif present in a sequence is oriented in structure. In the case of docked complex analysis, such things are seen.For analysis, different programs available show atomic interaction some programs even tell bond lengths. It has to be seen if the functional motif is correctly located or not. This is direct regulation i.e the amino acid sequence coding via DNA to form a protein. This regulation is not enough.

Indirect regulation, the environment affects/regulates protein function. (A biochemical term but in the perspective of molecular biology i is indirect genetic regulation). Biosynthesis of a catalyst is also regulated by certain enzymes this is indirect regulation. For example, prokaryotes synthesize their every requirement e.g lipid, etc for which they need enzymatic machinery. These enzymes have also encoded from a gene. Similarly, if one protein is in it then this indirect change is directly related to functional change.
Co & Post Translational Modifications:
- Ceratin PTMs are involved in protein native structure.
- In co-translational modifications, the translation process has not completed yet but modifications occur along with.

There is also a school of thought which says that such modification is also posted as that part has been translated but other schools of thought say it to be co.
Some post-translational modifications are permanent, some are temporary modification is glycosylation i.e. addition of sugars on protein.
•N-linked •O-linked
•NH2 acceptor •OH group
•Asparagine (Asn N) •Mainly serine & threonine
•NH2 acceptor •OH group
•Asparagine (Asn N) •Mainly serine & threonine
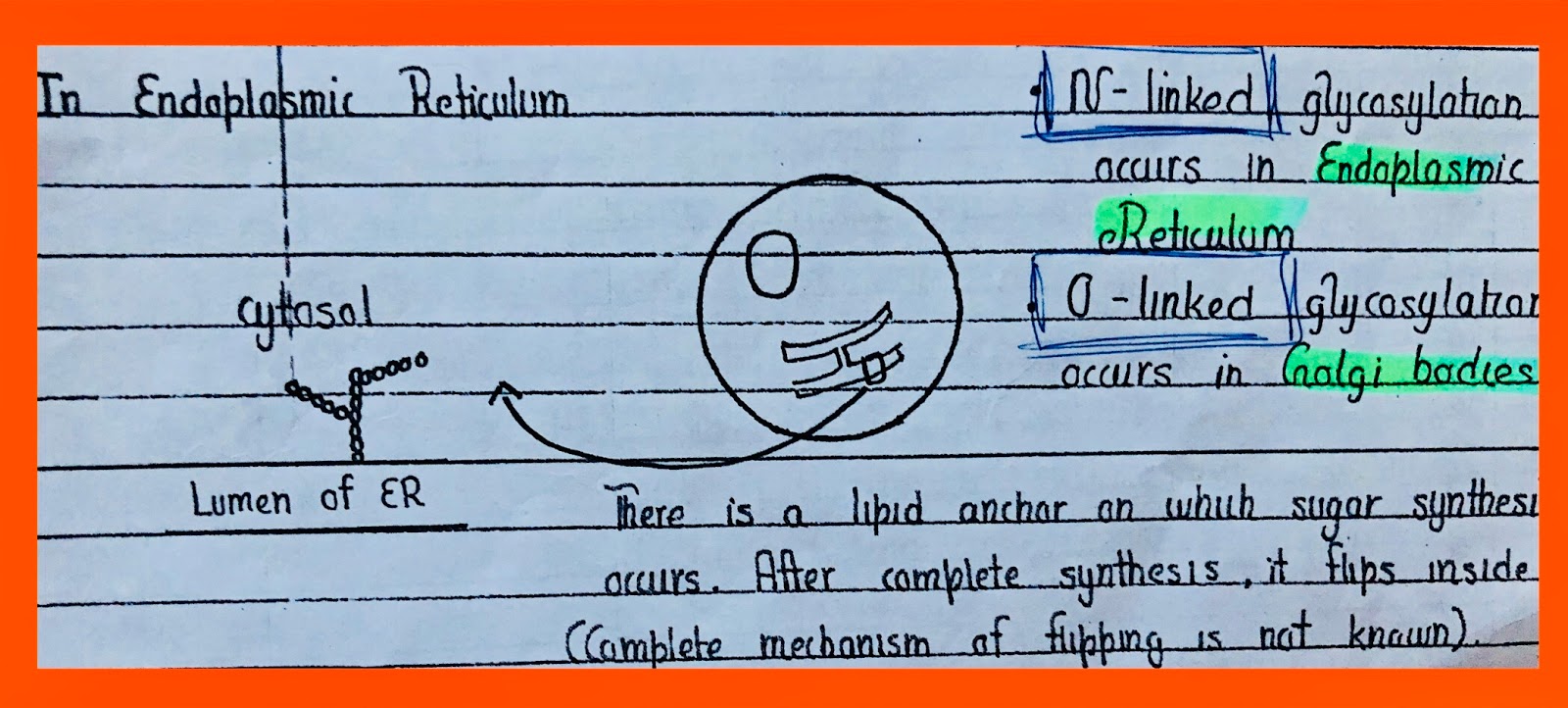
The sugar after flipping inside is removed and is then located on asparagine.
N-linked is one of those PTMs that are involved in stabilizing the 3-D structure of the majority of proteins. The endoplasmic reticulum is the start of the secretory pathway. Such proteins come in εR whose modifications are important to be carried. εR and Golgi bodies are processing machinery of the cell. Such proteins that either act as a receptor on the surface or secreted outside are processed here. In some proteins, both types of glycosylation occur.
On receptor/surface proteins and secretory proteins have a number of sugars present. This can be explained by the fact that the majority of epitope recognition sites are made by sugars (can be amino acids as well in some cases). The recognition memory of sugars is 100-1000 times more than proteins hence sugars by nature were chosen for this. Therefore, surface proteins and anchored proteins having the outside membranes have more sugars.
The chain of sugars that is synthesized is also processed. After flipping inside and before transferring to specific protein it is modified i.e trimming of some terminal sugars addition of some others. After transfer, protein folds. Proofreading also occurs here like in replication. Chaperones evaluate protein folding. Check the sugar contents by binding to terminal sugars. Where folding is wrong they change the sugars. When folding is correct the protein goes into Golgi and further modification is O-linked. The O-linked glycosylation sugars are added one by one, unlike N-linked where the whole chain is added.
Glycosyltransferases are present here:

Terminal Sugars
These are involved in recognization by changing them recognization proteins change.
It is important to know what type of sugars are present what are the sugars net to it and what type of linkage it is all these factors are important.
The purpose of the vesicle is to protect proteins from different interacting proteins.
- Another modification that occurs in the εR is Acetylation.
- For Methylation, methyltransferases are present.
- Phosphorylation & Nitrosylation are temporary/reversible.
- Ubiquitination & Sumoylation (ubiquitin & sumo) are permanent they do not remove until the protein is degraded. These modifications have a relation with the stability of the protein (Increase the stability)
- In εR & Golgi, enzymes are mostly membrane-anchored.
- In cytoplasm enzymes are free.
O linked glycosylation can occur in cytoplasm nucleopore complex & to some extent in nucleoplasm etc. Some of the O-linked glycosylation are permanent while some are temporary.
- Those that occur in Golgi are permanent.
- Those in the cytoplasm are temporary (only of single sugar molecules).
Editor's Recommendation:
- Analysing Metabolic Pathways
- Protein Threading Sequence
- Ab Initio Protein Structure Prediction
- Homology Modeling
- Hot Start PCR, Multiplex PCR, Avoiding Contamination In PCR, Advantages, and Disadvantages in PCR
- DNA Damage
- Docking | Protein-Protein Docking | Protein-Ligand Docking
- Functional Regulation | Genetic Aspect | Indirect Aspects
- Database Development
- PTMs and Functional Regulations
- Modeling Cellular Processes
- PCR Reagents | Stochastic Effect | STR Classification
- DNA Degradation
- DNA Quantification | Human DNA Quantification Method | Advantages
- Desirable Characteristics of STR used in Forensic DNA typing
- DNA Ladders
- Metabolic Pathways
- Non-Human DNA
- Mitochondrial DNA
- Integrated Genomic Circuits
- Shutter Product Formation
- STR Sites
- Mini STR Sites
- Molecular Diagnosis of Genetic Diseases
- Immuno Quantitative Assay
Functional Analysis At Structure Level
 Reviewed by Abdullah
on
June 17, 2020
Rating:
Reviewed by Abdullah
on
June 17, 2020
Rating:
Reviewed by Abdullah
on
June 17, 2020
Rating:
No comments:
Don't add any Spam link in comment box.